☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
superenrrollamiento del DNA
GENETICA VETERINARIA
Replicación
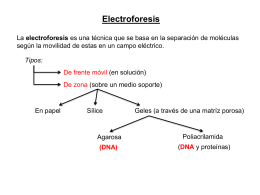
Clase Electroforesis
Transferencia de material genético II
Lab5_Enzimas Restriccion_UPRAg
Estimada(o) colega:
Biología Molecular Básica para el Patólogo
KARP_7a_c10_NATURALEZA_GENES_GENOMA
Las baldosas mojadas
Slide 1
Leyes Gases
Lección 5 : - Cálculo de estructuras y construcción
Gutiérrez Reynoso - Sociedad Argentina de Genética
puentes y bases nitrogenadas
2a CLASE SONDAS Y PCR
TEMA 1 Estudios de los esfuerzos y deformaciones en la región
Tema V Cilindros de pared gruesa