☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
KARP_7a_c10_NATURALEZA_GENES_GENOMA
Lo último en Genética: ¿Cómo lo afecta a Usted?
a un medio caliente.
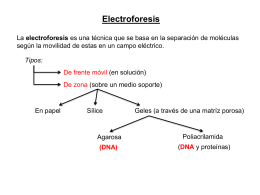
Clase Electroforesis
Replicación
superenrrollamiento del DNA
ALCANCES DEL PROYECTO GENOMA HUMANO
Transferencia de material genético II
caracteristicas_de_los_seres_vivos
Genoma Humano - Blog de Química Biológica Patológica
Lab5_Enzimas Restriccion_UPRAg
SE NECESITA AYUDANTE
Tipos_de_celulas
Los editores y el libro digital
presentación Power Point - Universidad de Valladolid