☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
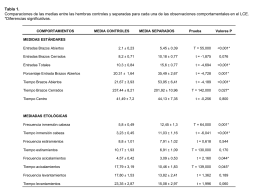
No category
Macroevolucion_Hot_version_extendida_PT_2015
El lobo marino sudamericano
CIERVO
Sistemas de apareamiento
Descarga
1 Evolución de los primates
Microsoft PowerPoint
Ver Programa
Genética General Laboratorio - Instituto de Investigaciones
sistemas de apareamiento
resultados
Modelos y Machos
Consecuencias de la selección sexual
Conceptos de apoyo para participar en el seminario
Insectos
una amiga que está estudiando biología en Una Universidad