☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
Presentación de PowerPoint
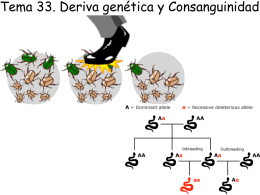
TEORIA NEUTRAL DE LA EVOLUCION MOLECULAR
INTRODUCCION
Tema33-w0910
DATOS Y AZAR - Colegio SIS Quillota
Estadística y Probabilidad I
EN BUSQUEDA DE LA INMORTALIDAD
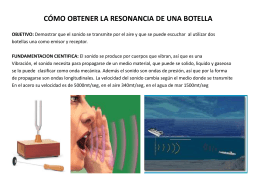
cómo obtener la resonancia de una botella objetivo
Análisis de varianza
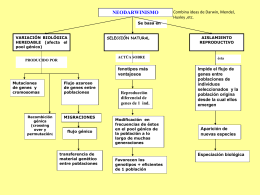
Genética de POBLACIONES
evolucion_parte_ii
Guillermo Garberi - Viernes 06 de junio
Genética de Poblaciones
Carro eléctrico casero.
Evolución y Especiación