☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
INTRODUCCION
Diapositiva 1 - Inicio | Universidad Virtual de Salud
Diapositiva 1
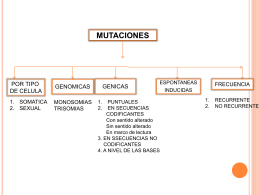
mutaciones
Presentación de PowerPoint
Control del Comportamiento
Diapositiva 1
Diapositiva 1
DATOS Y AZAR - Colegio SIS Quillota
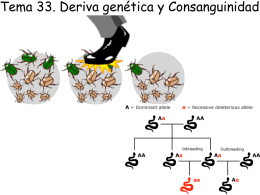
Tema33-w0910
Genética de POBLACIONES