☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
COMUNICACIÓN CELULAR
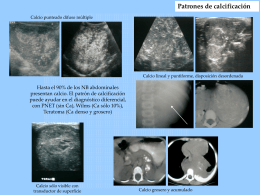
4. Patrones de calcificación
LIGANDOS Y RECEPTORES
segundos mensajeros (final)
Spay Oklahoma - Calcium Chloride Castration
MEDICACION, TRATAMIENTO FARMACOLOGICO Y OBTURACION
UN CAMBIO DE VIDA UN SISTEMA UNICO
Nomenclatura y notación química de las sales ternarias
Transparencias
Problema "La mala alimentación"_Julio Tellez
Presentación de PowerPoint
Alimentos cardiosaludables