☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
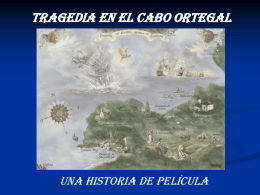
No category
Power Point
ver Power Point
IMPACTO DEL EFECTO ALLEE EN LAS PRESAS EN LA DINÁMICA
presa
1 EL ESTUDIO DEL MEDIO AMBIENTE
ver Power Point
Presentación de PowerPoint
Juana Leslie Aguilar Bernal
Centrales
Matrices Random de Leslie
Vuelo de Ibis
Presentación de PowerPoint - ESTUDIANTES NOCTURNOS
Tercer hábito
Diseño de presas
Tercer Habito, Nydia Mendiola A, 2005-2