☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
Diapositiva 1 - WordPress.com
Las bacterias como fábricas de proteínas
Transcripción del ADN. El ARN mensajero. Traducción genética
origen de la vida
Síntesis de Proteínas
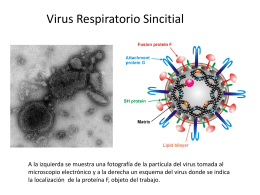
Virus-Respiratorio
Power Point 3 - biomilenio.net
DESNATURALIZACIÓN DE LAS PROTEÍNAS GLOBULARES
Proteína - Bioquímica para Enfermería
¿Por qué pide el mercado memorias de traducción?
RNA mensajero en eucariotas