☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
Diapositiva 1 - OpenCourseWare de la Universidad de …
potencial de membrana
TRANSPORTE ACTIVO CELULAR - Biología 100 -
Soluciones
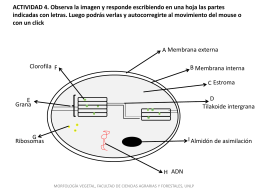
Diapositiva 1 - Morfología Vegetal
Diapositiva 1
Diapositiva 1
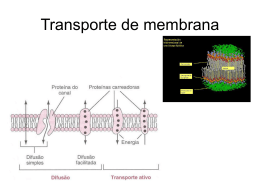
Mecanismos de transporte - BluWiki
Diapositiva 1 - art3vespenmovimiento
UNIVERSIDAD INTERAMERICANA DE PUERTO RICO Recinto de
Pricast
Enlaces químicos