☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
Capitu
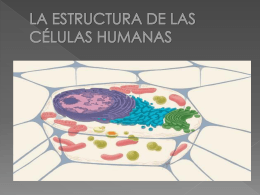
3-La estructura de las células humanas (El trío tralará)
Diapositiva 1 - art3vespenmovimiento
Biología Articulo Tercero Constitucional
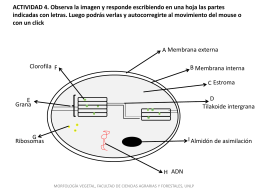
Diapositiva 1 - Morfología Vegetal
Soluciones
Diapositiva 1
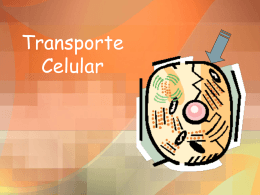
Cell Transport
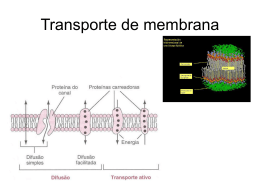
La membrana y el transporte activo
Presentación de LA CÉLULA
Presentación de PowerPoint