☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
Diapositiva 1
Tema 14
donde quedo la bolita
METABOLISMO DEL PIRUVATO - Profesora Maribel Arnes
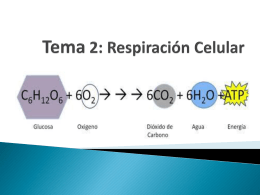
Respiracion
TRANSPORTE ELECTRONICO Y FOSFORILACION OXIDATIVA
Document
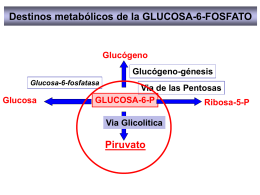
CATABOLISMO DE LA GLUCOSA
METABOLISMO - quimicabiologicaunsl
File