☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
Document
Lecture 1.1 Introduction to the Course … and Computers
Syllabus - faculty at Chemeketa
Syllabus - faculty at Chemeketa
Document
A Highly Reliable Fault-tolerant Airline Reservation System
Mecanismos de alta disponibilidad con Microsoft SQL …
History and Impact of Sci. Vis.
Introduction to Organic Chemistry 2 ed William H. Brown
Writing science in plain English
Genetic Genealogy, History and Prehistory, and DNA
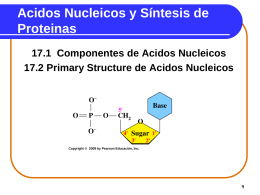
Nucleic Acids
No Slide Title
Mutation Testing
Welcome to Chem 152 - San Diego Mesa College
Service Oriented Architectures in Heterogeneous …
Microbiology
Error Control Coding, Cancer Development and Statistical
Finding Inconsistencies using Relation Partition
Document
GENE to PROTEIN - Jefferson County Public Schools
SWE 637: Syntax
Focused Reusable Exercises for Teaching Effective Test …