☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
No category
Equilibrio de Hardy-Weinberg y sus desviaciones
DATOS Y AZAR - Colegio SIS Quillota
FORMAS DE REPRESENTAR LAS
Presentación de PowerPoint
Distribución de frecuencias
Clase4_JJF_2015
Variables cualitativas y cuantitativas
Descarga - Profe Marcelo.com
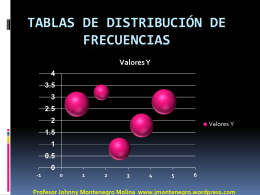
Tablas de distribución de Frecuencias
Presentación PPT - Freddy Villabona C.
Presentación de PowerPoint
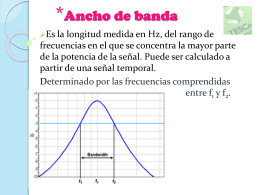
Ancho de banda - 7421MARZO
Tema 2: Patrón de apareamientos y equilibrio Hardy