☰
Explorar
Iniciar sesión
Crear una nueva cuenta
Pubblicare
×
Descargar
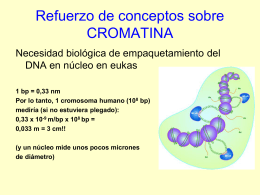
No category
histonas
EL MATERIAL GENETICO
ME DE CROMATINA EXTRAÍDA EN LA FORMA EXTENDIDA Y
El núcleo
IMPRONTA GENOMICA (“IMPRINTING”)
Diapositiva 1 - LGVH
Presentación en power point
CROMOSOMAS ANILLO
sólo quedan 5 semanas ingenieria genetica
Lo que se hereda no se roba no?? O será mentira?
Mitosis y Meiosis
Laboratorio Enzimas de Restriccion II
ADN - campvs.cl
t.6. nucleotidos y acidos nucleicos
EL NÚCLEO.
Translational and rotational settings of H2A.Z